
Images - Arranged by Taxonomic Groups
| Angiosperms | Bryophytes | Caytoniales | Conifers | Cycadophytes | Ferns | Ginkgophytes | Leptostrobaleans |
|---|
| Lycophytes | Sphenophytes |
|---|
Cycadophytes |
Arctic Fossil Cycadophyte Images |
The presence of cycad foliage, and in particular the common occurrence of Nilssonia leaves (most notably N. serotina, N. alaskana and N. yukonensis) at high paleolatitudes in the Cretaceous of Northeastern Russia and Northern Alaska has been used as evidence for frost free, warm temperate, if not subtropical, near-polar conditions (Smiley, 1967) and contributed to the concept of the Cretaceous being a time of extreme global "greenhouse warmth". Underlying this interpretation was the assumption that the Cretaceous cycads had a similar biology and climatic tolerance to their living relictual relative which are restricted to subtropical and tropical climates even though a few can resist light frosts. This assumption was made despite the fact that no manoxylic bulbous trunks, typical of many living cycads, had ever been found as Arctic fossils (Vakhrameev, 1991).
Nilssoniocladus and the question of obliquityThe genus Nilssoniocladus was first described in 1975 by Kimura and Sekido based on material from the early Lower Cretaceous Oguchi Formation, Itoshiro Group in the Tetori Basin, Central Honshu, Japan. The type material, Nilssoniocladus nipponensis represents a plant with a thin vine-like stem bearing spirally arranged dwarf short shoots with persistent leaf scars. The leaves, assigned to Nilssonia nipponensis Yokoyama, were borne in clusters of three to seven or more at the distal ends of the dwarf shoots. Although Nilssoniocladus was assigned by Krassilov (1987) to its own order it is still widely regarded as representing an extinct member of the Cycadales. Kimura and Sekido considered Nilssoniocladus to be deciduous and the thin vine-like stem was likely to have been more frost resistant than the pachycaulous trunks of modern evergreen thermophyllic cycads. Spicer (1987), Parrish and Spicer (1988a), Spicer et al., (1992); Spicer and Chapman (1990) used the Nilssoniocladus model for polar cycads on the assumption that Nilssonia leaves in Alaska represented the same type of plant as that found in Japan. Taphonomic evidence involving a wide range of foliage forms (including conifers, angiosperms, ginkgophytes and cycadophytes) was used to argue for wholesale deciduousness in the Cretaceous polar forests (Spicer and Parrish, 1986; Spicer, 1987; Parrish and Spicer, 1988a; Herman, 1994). However it was not until until Spicer and Herman (1996) discovered Nilssoniocladus shoot material in both the latest Albian Killik Tongue of the Chandler Formation, N. Alaska, and the Cenomanian Grebenka locality in Russia, that this assumption was validated. |
 |
|
|---|---|---|
Modern cycads pictured above are susceptible to freezing conditions and are therefore restricted to sub-tropical and tropical conditions, the only exceptions being Bowenia that can survive frosts by having underground stems. The manoxylic stems have little woody tissue in them and are particularly vulnerable. The orange and yellow structures at the top of the trunks are reproductive structures. |
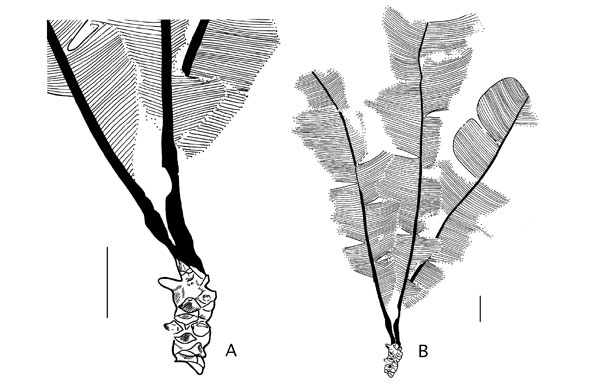
Nilssoniocladus is the name given to one or more Nilssonia leaves attached to a short shoot; an ensemble that was clearly shed from the parent plant as a discreet unit or 'rosette' of leaves. These are illustrated on the right. The leaves associated with the Alaskan stems are referable to Nilssonia alaskana Hollick (1930). They are characterised by well dissected laminae, the sinuses between leaf segments or lobes being rounded and the apices also rounded. The spacing between segments is approximately the same as the width of the segments and the more distal segments on each leaf tend to be apically curved. |
 |
|
|---|---|---|
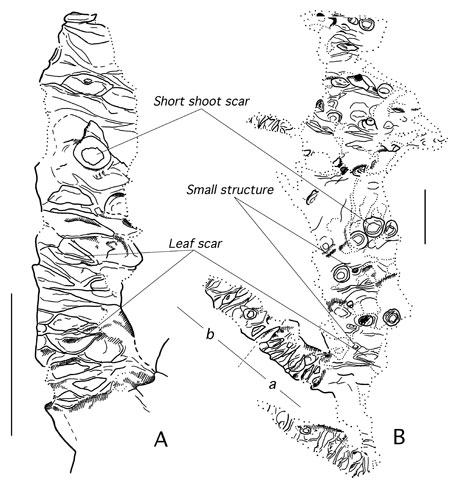
In addition to leaves attached to short shoots and discarded from the parent plant as the Nilssoniocladus unit, Spicer and Herman found elongated stems with short shoots identical to those of Nilssoniocladus, togther with Nilssonia leaves. The constant morphology of the short shoot scars was regarded by Spicer and Herman (1996) as strong evidence for an abscission layer, which coupled with the frequent leaf 'rosettes' attached to a short shoot that was clearly shed as a separate unit, provided strong evidence for shoot shedding, as well as individual leaf shedding, in Nilssoniocladus. The frequency of short shoot scars and their co-occurrence with short shoots in place is evidence that there were many separate cohorts of short shoots produced throughout the life of the plant. This illustrated in the drawing to the right. The stem of Nilssoniocladus was evidently robust and long lived. The presence of at least two distinct and complete growth increments, and the partial presence of two others at each end of the stem in the holotype of N. alaskensis (right), shows that this individual probably grew for a number of years. Judging by the lack of taper of the stem it is possible that the plant could have lived well in excess of ten years. If the portion of stem preserved represents a lateral branch of a larger plant the lifespan could have been considerably longer. The variability in the length of the growth increments probably reflects climatic variations from year to year. These growth increments were interpreted by Spicer and Herman (1996) as being produced annually. For images of the actual specimens reported by Spicer and Herman (1996) click here. |
 |
|---|---|
| Long shoot stems associated with Nilssoniocladus alaskensis Spicer and Herman showing short shoots and abcission leaf scars where the leaf 'rosettes' were attached. Scale bars represent 1cm. (From Spicer and Herman, 1996). |
The discovery of Nilssoniocladus and stems with short shoots identical to those at the bases of Nilssoniocladus 'rosettes', coupled with leaf mats suggesting Nilssonia leaves were shed synchronously, is powerful evidence that the Nilssonia/Nilssoniocladus plant was deciduous and not evergreen. This is significant because the presence of cycads, the only living examples of which are thermiophilous evergreen plants, had been used not only as evidence that the Arctic was once warm, but that the Earth's obliquity had also changed significantly (Douglas and Williams, 1982). Because Nilssonia leaves are known to have lived at paleolatitudes of 75 °N or more and if the obliquity of the Earth was the same as present the source plants must have experienced continuous winter darkness of at least six weeks, bounded by a continuous twilight period of three weeks in the spring and autumn (Anonymous, 1978). A warm temperate to subtropical frost-free climate, combined with this light regime and evergreenness in which a living leaf load was borne throughout the year, would, it was argued, have led inevitably to a high respiratory drain on the plant's resources during the winter: young plants in particular would have been vulnerable. A reduced obliquity, however, would have reduced the duration of winter darkness and evergreen plants might have been able to survive in near polar environment (Douglas and Williams, 1982; Wolfe, 1978). With the discovery that the Nilssonia/Nilssoniocladus parent plant was deciduous and had a woody vine like stem tolerant of frost meant that reduced obliquity was no longer required to explain the presence of cycads at high latitudes. Leaf shedding and dormancy during the polar winter darkness would have ensured survival through this critical season.